Survivors don't need an overview, they need EXACT PROTOCOLS THAT DELIVER RECOVERY. This doesn't do that; USELESS!
I'd have everyone associated with this fired, include mentors and senior researchers proposing or approving any part of this!
An overview of the effects and mechanisms of transcranial stimulation frequency on motor learning
Journal of NeuroEngineering and Rehabilitation volume 21, Article number: 157 (2024)
Abstract
Many studies over the recent decades have attempted the modulation of motor learning using brain stimulation. Alternating currents allow for researchers not only to electrically stimulate the brain, but to further investigate the effects of specific frequencies, in and beyond the context of their endogenous associations. Transcranial alternating current stimulation (tACS) has therefore been used during motor learning to modulate aspects of acquisition, consolidation and performance of a learned motor skill. Despite numerous reviews on the effects of tACS, and its role in motor learning, there are few studies which synthesize the numerous frequencies and their respective theoretical mechanisms as they relate to motor and perceptual processes. Here we provide a short overview of the main stimulation frequencies used in motor learning modulation (e.g., alpha, beta, and gamma), and discuss the effect and proposed mechanisms of these studies. We summarize with the current state of the field, the effectiveness and variability in motor learning modulation, and novel mechanistic proposals from other fields.
Introduction
The use of transcranial alternating current stimulation (tACS) has grown in popularity in recent years as it allows researchers to study the effect of electrical stimulation on the brain at specific frequencies. At present, the exact neurophysiological mechanisms behind tACS remain elusive, however the literature on tACS can provide insights into new approaches to target motor deficits and/or enhance motor performance. Here we review major works in the field of tACS and motor learning in order to recover insights surrounding how tACS may alter neural oscillations, the differences in methodological approaches, and the different task-stimulation pairings which were most and least effective in finding effect. This literature review intends to serve as a pause in the field to recognize the most promising, and more contentious, advancements.
Motor learning is intrinsic to all humans and animals and involves the acquisition of a new skill followed by consolidation. Acquisition of a motor skill is associated with various cortical and subcortical brain regions, however the primary motor cortex (M1) is often the most discussed key region in motor learning [1]. Krakauer et al. [1] describes motor learning as bipartite as learning a motor skill not only requires skill acquisition, but also skill maintenance, which is just as important as acquisition due in part, to a changing environment. The inevitable onset of ageing essentially forces the individual to modify an already learned skill in order to adapt to a decline in motor competency. With an increasingly ageing population, neuroscientists and psychologists have been trying to gain a deeper understanding of how the manipulation of motor learning may have clinical implications. A plethora of studies in the last two decades have assessed the effect of transcranial electrical stimulation (TES) on motor learning to investigate whether application of a non-invasive weak electrical current to the scalp can modulate neuronal excitability and subsequent motor learning. The first human study that successfully induced excitability in the motor cortex through transcranial direct current stimulation (tDCS) was demonstrated by Nitsche and Paulus [2]. Two electrodes were attached to the scalp of the participant and a current applied at a constant rate over time. It was found that anodal current applied to the scalp resulted in cortical excitability and subsequent neuronal depolarisation, and it was suggested that these effects were due to a shift in the neuronal resting membrane potential. Up to this point, only invasive techniques had successfully produced intracranial currents. Zaehle et al. [3] demonstrated the first electrophysiological evidence of modulation of rhythmic brain activity by application of transcranial alternating current stimulation (tACS) to the occipital cortex. TACS differs from tDCS as it involves the application of a sinusoidal waveform at a specific frequency which alternates between the anode and the cathode as described by Pollok et al. [4]. Zaehle et al. [3] demonstrated that application of tACS to the occipital lobe at the individuals alpha frequency (iAF) resulted in amplification of this frequency following 10 min of stimulation, and Helfrich et al. [5] expanded on this the point, that exogenous stimulation of the brain over the parieto-occipital region can synchronise with cortical oscillations endogenously. The suggestion that endogenous brain oscillations could be entrained to a specific frequency by employing tACS paved the way for future research in the modulation of brain oscillations in a frequency-dependent manner. Later studies found that tACS at specific frequencies could alter motor movement as demonstrated by Pogosyan et al. [6]. More recent literature has linked tACS with motor learning [4] however the most effective frequency stimulation for modulation of motor learning has been the topic of immense debate and numerous attempts.
The present review aims to address this issue by considering from the recent few decades which individually investigate different stimulation frequencies. Much of the literature involves the application of tACS to the motor cortex as this region of the brain is largely known to be involved with motor control, however this is not always the approach. This review will provide an overview of the main tACS frequencies investigated throughout the literature to alter motor learning including, theta, alpha, beta and gamma frequencies, gathered from recent and what we believe to be influential works available online and in English. Similar reviews such as that by Takeuchi and Izumi [7] discuss the effects of tACS on brain activity, and so this review will focus more specifically on frequency specific effects in motor learning. This will include general proposed mechanisms for each approach, and future recommendations for motor learning modulation with tACS and its clinical relevance will be discussed.
Theta (4–8 Hz)
Beginning with slow-wave tACS, theta stimulation has had arguably the most straight-forward effect on motor learning compared to other frequency bands.
Alpha (8–12 Hz)
In one of the earliest examples of alpha stimulation for motor learning, Antal et al. [8] assessed the effects of stimulation on cortical excitability in humans using alpha-tACS delivered to the motor cortex. They employed the serial reaction time task (SRTT) paradigm, which has been used extensively to assess learning of a repeated motor sequence with decreased reaction time an indicator of enhanced motor learning. A 7-min stimulation of 10 Hz tACS over the motor cortex was found to improve implicit learning, as evidenced by a faster decrease in reaction time at 10 Hz tACS compared to sham stimulation, however no significant changes were observed in the EEG, nor with motor evoked potentials.
It was suggested by Helfrich et al. [5] that human oscillatory brain activity could be entrained by tACS at the parieto-occipital cortex, which is thought to play a role in proximal and distal movements particularly for prehensile purposes [9], although its role in motor learning is less clear. EEG recordings show that alpha power was enhanced in the parieto-occipital cortex with 10 Hz tACS, and an additional target detection task showed that participant performance was enhance. It was thus postulated that stimulation of alpha neuronal oscillations leads to alpha-band synchronisation and may increase the ability to distinguish between both relevant and irrelevant sensory information required for a particular task thus improving the efficiency of brain components required for target detection. Hence, phase-dependent enhancement of target detection performance through exogenously stimulating alpha activity may contribute to motor learning, however we highlight that this influence is more indirect. Furthermore, motor learning was found to correlate positively with implicit motor learning, albeit there is little evidence to support alpha power changes as a marker of sequence learning, but rather as a marker of attention control [10, 11].
According to Pollok et al. [4, 12], motor learning is associated with changes in primary motor cortex oscillations. It is suggested that motor control is frequency-dependent, such that alpha oscillatory synchrony may be present during automatic motor control, whereas beta oscillatory synchrony may be present during explicit motor control and potentially motor sequence learning. This was made experimentally clear in a study by Pollok et al. [4] whereby both alpha (10 Hz) and beta (20 Hz) tACS was applied to the left M1, and it was found that 10 Hz and 20 Hz tACS significantly improved learning of the task as observed by shorter reaction times for both, compared to 35 Hz tACS and sham stimulation. Although it is proposed that both alpha and beta frequency stimulation assist motor learning, the interference introduced by the random sequence was shown to decrease the effectiveness of 10 Hz stimulation while 20 Hz stimulation remained unaffected, suggesting that alpha frequency stimulation is effective for the initial aspect of motor learning but does not contribute to the maintenance of a learned sequence and subsequent motor consolidation as observed with beta stimulation. Another study addresses the issue of motor consolidation decline in older adults [13]. The M1 was stimulated offline with 10 Hz and 20 Hz tACS immediately after training for 15 min and motor learning was assessed by speed and accuracy of sequence learning six hours post stimulation. The main finding suggests that 10 Hz stimulation seems to reduce motor consolidation compared to sham stimulation as seen by an impaired retest performance, positing that alpha-tACS applied to the M1 potentially interferes with the offline processing of a learned motor sequence in older adults. Increased alpha power has been associated with decreased cortical excitability via alpha wave inhibition, and an obvious explanation for the above findings would be that endogenous alpha oscillatory brain activity enhanced by tACS is associated with increased cortical inhibition in the M1. However as Fresnoza et al. [14] found that alpha-tACS on the M1 improved motor skill consolidation in older people, Rumpf et al. [13] interpreted their findings differently, arguing that local modulation of alpha oscillatory activity in the M1 may alter network synchrony between different brain networks connected to the M1. Disruption of communicative networks by alpha oscillation modulation may contribute to impaired consolidation or worsen already impaired consolidation in older people. In one study, the causal link between impaired alpha activity and motor impairment was tested in ageing individuals. Fresnoza et al. [14] delivered individual alpha peak frequency (iAPF) stimulation and iAPF + 2 Hz to the left motor cortex of individuals during a standard SRTT. A 5 min resting state EEG was recorded for each participant prior to stimulation to identify their iAPF. Findings demonstrated that tACS applied at both iAPF and iAPF + 2 improved motor consolidation in older, but not younger participants. Although the authors note the possibility of a ceiling effect that might explain a lack of improvement following stimulation, it is suggested that effectiveness is related to the synchronization of impaired endogenous alpha waves in the older population. While modulation in young people may have decreased cortical inhibition as younger individuals did not have impairment in alpha oscillatory activity prior to stimulation. Therefore, it is postulated that modulation may have different effects depending on the nature of endogenous oscillatory activity within an individual. It is suggested that the neurophysiological after-effects of alpha-tACS may be induced by individual alpha peak frequency and not by fixed alpha frequency stimulation, which supports the findings of Fresnoza et al. [14] above who argued that iAF tACS can induce neurophysiological aftereffects and subsequently generate changes in oscillatory activity that are concomitant with motor consolidation. Hence, fixed alpha-tACS may facilitate initial motor learning even when applied in offline conditions, however motor consolidation and the maintenance of a learned motor skill may be induced by tACS at iAF rather than at a fixed alpha frequency. Harada et al. [15] demonstrated that 10 Hz tACS stimulation post-visuomotor learning facilitates angular learning correction compared to 20 Hz stimulation and sham stimulation although, critically, no significant differences were observed in oscillatory neural activities between 10 Hz, 20 Hz and sham offline stimulation.
There has been speculation that alpha neural oscillations are involved in communication between brain regions which was explored by Schubert et al. [16] who found that 10 Hz tACS to the right cerebellum (rCB) interfered with learning in the SRTT compared to sham stimulation, yet this was not observed following 10 Hz tACS application to the left primary motor cortex (lM1) compared to sham stimulation. Since the cerebellum is associated with motor planning [17] and error correction [18], it is speculated that the cerebellum may contribute to motor learning under higher frequency oscillations such as gamma frequency rather than slow frequency oscillations such as alpha frequency. This was in fact investigated by Miyaguchi et al. [19] and will be discussed later. It could further be speculated that stimulation at non-endogenous frequencies (i.e., delivering alpha tACS to the more gamma-oriented cerebellar frequencies) might resulting in slowing or disruption of these signals, and further impairment of performance, or further that specifically alpha stimulation (an inhibitory signal) simply disrupts regional processes [20]. Here we also highlight the importance of location when delivering tACS, since delivery of alpha stimulation has also shown to differ in effect between M1 or cerebellar montages.
Findings showed that alpha coherence between the premotor cortex (PMC) and the rCB was enhanced following rCB tACS, and correlated with motor learning, indicating that alpha oscillations applied exogenously and induced endogenously in the rCb may mediate information transfer in a premotor–cerebellar loop during the learning of a motor sequence [16]. This therefore may be a mechanistic pathway by which alpha oscillatory activity contributes to initial motor learning. Conclusively, the use of alpha-tACS for motor learning appears to be effective when modulating networks with existing alpha band dynamics, as evidenced by iAF stimulation effectiveness, but is disruptive when delivered to regions which would under normal motor learning dynamics not depend or improve with inhibitory effects.
Beta (13–30 Hz)
Canonical association between beta and motor function implicates beta-tACS as a common target within the field. Feurra et al. [21] describes beta oscillations as “idling” beta activity in human sensorimotor regions and aimed to understand the association of beta “idling” activity with human corticospinal output. The size of motor evoked potentials (MEP) was found to increase with beta-tACS (20 Hz) only, suggesting that idling beta rhythm of sensorimotor areas facilitates corticospinal neuron firing and increases cortico-muscular coherence, an important aspect in the context of motor learning [22].
Pollok et al. [4] applied 20 Hz tACS during an SRTT motor learning task to the M1 of young individuals and found that initial acquisition was increased compared to sham stimulation. Although Antal et al. [8] previously found that 15 Hz tACS did not facilitate motor learning, they argue that 15 Hz does not match beta oscillatory activity and fails to entrain beta brain oscillations, similar to the effectiveness of iAF, and consistent with the pursual of personalized tACS. Furthermore, varying methodological approaches were used by Antal et al. [8] thus the two cannot be compared. Findings from Pollok et al. [4] showed that 20 Hz tACS was concomitant with less susceptibility to an interfering random pattern in SRTT compared to 10 Hz tACS, and was said to favour stabilization of a learned sequence, suggesting that beta neuronal oscillatory activity may contribute to maintenance of a learned sequence and early consolidation. Moreover, 20 Hz tACS may also contribute to the processing of the association between a visual cue and a button press [23].
In a study by Krause et al. [24], the possibility of differentially affecting the retrieval of a newly learned sequence by tACS at varying frequencies was investigated with the hypothesis that modulation of beta oscillations with tACS is a prerequisite for functional reorganisation associated with early consolidation of learned motor skill. tACS was applied at 10 Hz, 20 Hz and sham stimulation and learning was assessed with a standard SRTT where reaction times indicated learning ability. While no significance difference in reaction times was observed between the three stimulation groups at the end of acquisition, reaction times were significantly different between stimulation groups during retrieval of the learned sequence. Reaction times did not improve after a period of reacquisition for both 10 and 20 Hz tACS suggesting that 20 Hz tACS is more useful to facilitate learning with less training rather than further training. To further support the findings above, Pollok et al. [12] found that beta power suppression during acquisition of a motor sequence was positively correlated with improvement of reaction times in a sequential learning task, in line with the hypothesis that beta activity represents a marker of reorganization during sequence learning [25]. Despite the lack of neurophysiological evidence, it is speculated that beta neuronal oscillatory modulation may encourage consolidation of a motor skill through functional reorganisation of the M1 and subsequent maintenance of a learned sequence with Pollok et al. [12] further suggesting that changes in beta and gamma oscillatory power are involved in motor learning.
Bologna et al. [26] tested the effects of beta-tACS and gamma-tACS on the M1 during motor learning by assessing the speed at which an individual abducted their index finger in response to a “Go” signal. Two findings support the argument that beta-tACS may disrupt movement execution: (1) A lack of improvement in finger speed compared to sham and gamma stimulation was observed indicating reduced motor learning and (2) Cortical excitability in the M1 was not altered by beta-tACS, ruling out an effect of tACS on cortical excitability. It should be noted however that task pairing in that studied differed from previous sequence learning tasks, which raises questions concerning the role of task demand and circuit engagement and observed tACS effects. Pogosyan et al. [6] have shown that boosting beta band cortical activity slows human movement, so an explanation for the findings of Bologna et al. [26] may be that beta-tACS has antikinetic effects on human movement which prevented an improvement in the motor learning task. Additionally, it is argued that, due to a lack of correlation between cortical excitability and beta-tACS, one cannot support the hypothesis that behavioural performance and cortical activity are synergistic [27].
Accordingly, beta-tACS may modulate motor learning via multiple mechanisms, either via the alteration of idling beta activity for cortico-muscular coherence [21, 28], or through synchronization of beta-rebound following motor response during learning [7]. It is however apparent, that beta tACS is ineffective at modulating cortical excitability in the motor areas, and further at transiently entraining oscillations [27, 29], and in fact beta tACS is theoretically an ineffective modulator of consolidation due to the dynamics of reduced beta power [26], cortical reorganization [25], and eventual consolidation [30].
Gamma (30–80 Hz)
Sugata et al. [31] investigated whether tACS at varying frequencies could alter oscillatory neural activity and increase the capacity for motor learning. Prior to this study, little research had investigated motor learning capacity, with the focus of much of the literature pointed towards motor learning acquisition and consolidation, and the hypothesis was that tACS entrains oscillatory neural activity and subsequently increases motor learning capacity contributing to improved learning. A visual cue button test with decreased reaction time as an indicator of motor learning was used, and after application of 10 Hz, 20 Hz, 70 Hz and sham stimulation individually to the left M1 of participants, results showed that 70 Hz-tACS significantly decreased reaction time compared to sham stimulation. In support of this, MEG studies examined oscillatory neural activities and showed that 70 Hz tACS significantly increased beta-band power of the M1, likely via cross-frequency modulation. Although the mechanisms remain elusive, previous studies have reported cross-frequency coupling (CFC) [32], plays an important role in motor learning capacity and ultimately contributes to the aftereffects of tACS such as motor skill consolidation. Bologna et al. [26] found that gamma-tACS to the M1 induced a small improvement of finger abduction speed although a detrimental effect on motor retention compared to baseline conditions was observed indicating that gamma-tACS modulates initial motor learning rather than the retention phase of motor learning. There is indeed a scarcity of literature investigated the effect of slow-gamma band frequency in the M1 and its role in motor learning up until Giustiniani et al. [33] with the exception of Pollok et al. [4] who used slow gamma-tACS (35 Hz) as a control group in their previous study. Although no significant learning occurred with 35 Hz tACS compared to alpha and beta-tACS, the author did not rule out the possibility of a very weak effect of 35 Hz stimulation compared to the other two.
To investigate slow gamma-tACS, Giustiniani et al. [33] applied 40 Hz tACS over the M1 while participants performed a standard SRTT eliciting slower reaction times during 40 Hz tACS in the later sequential learning blocks, compared to sham stimulation and 1 Hz stimulation. It is postulated that slow gamma-tACS may disrupt cortical reorganisation required for retrieval of a learned sequence subsequently increasing susceptibility to interference induced by the random sequence of the SRTT. As earlier acquisition was not affected, and TMS-evoked MEP amplitudes were significantly reduced after 40 Hz tACS, it is suggested that 40 Hz tACS interferes with consolidation of a learned sequence by weakening the networks oscillating at beta frequency that are necessary for motor sequence consolidation. Hence, 40 Hz tACS may attenuate cortical reactivity required for the plastic changes concomitant with motor sequence consolidation. Additionally, previous studies suggest that high gamma-tACS improves motor learning through the activity of GABAergic interneuron activity [31] while slow gamma-tACS reflects glutamatergic excitatory pyramidal neuronal activity subsequently contributing to a reduction in motor learning competency [33]. It therefore appears that gamma tACS produces variable effects depending on low or high frequency range when delivered to the motor cortex, and further that these ranges produce differing physiological effect.
It is well established that the cerebellum is responsible for the modification of motor planning as well as the correction of motor errors, and the M1 receives input from the cerebellum forming a neural network that contributes significantly to the accuracy and smoothness of movement [34]. The “binding theory” describes the strengthening of neural networks through synchronisation of gamma band oscillation, and thus application of gamma-tACS could potentially modulate the activity of both the M1 and the cerebellum improving overall motor control as a result [19]. Considering this, they applied gamma-tACS and beta-tACS over the M1 and the cerebellar cortex region during the performance of a visual cue isometric force task. Only individuals receiving gamma-tACS, who initially displayed poor performance in the task, elicited an improvement in motor performance- and no improvement was observed in those receiving beta-tACS. Moreover, motor performance was only improved when gamma-tACS was applied simultaneously over both the M1 and the cerebellar hemisphere with a lack of improvement observed when gamma-tACS was applied individually. Taken together, the results of Miyaguchi et al. [19] support the “binding theory” phenomenon previously described which may contribute to improved motor performance. However, one cannot rule out the possibility of input from various brain regions that may very well contribute to the improved motor performance observed. In a later study by Miyaguchi et al. [34], motor learning was assessed with a visuomotor control task to investigate whether gamma-tACS of both the M1 and the cerebellar hemisphere influences motor learning by strengthening the neural network between the M1 and the cerebellum. Two separate groups received either sham or gamma-tACS while performing the motor task and results showed that gamma-tACS improved motor learning retention 24 h after the motor task compared to sham stimulation despite a lack of effect observed in motor learning efficiency. It is proposed that motor learning retention may have been improved through the strengthening of connections between the M1 and the cerebellar hemisphere and that cross-frequency coupling (CFC) may have occurred in the M1 by suppression of beta-oscillatory activity and enhancement of gamma-oscillatory activity, subsequently improving neuronal reorganisation associated with motor learning retention. Hence, further research is required to understand the role of beta oscillatory activity in various phases of motor learning as well as the interaction of beta oscillations with gamma frequency stimulation. Here again we also note the consideration of montage location, especially considering that inter-regional stimulation montages theoretically act different to single-site delivery, an aspect more extensively focused on in Takeuchi and Izumi [7].
Akkad et al. [35] describe phase amplitude coupling (PAC) of theta-gamma oscillations to be a fundamental process for cortical computation across the neocortex as it is already well distinguished that hippocampal activity such as memory encoding is modulated by coupling of theta-gamma oscillations. Thus, Akkad et al. [35] investigated whether theta-gamma coupling in the neocortex also plays a role in motor skill learning by applying 75 Hz tACS to the M1 at a 6 Hz theta peak during a thumb abduction learning task. Findings showed enhanced thumb abduction acceleration with theta-gamma tACS compared to sham stimulation which was particularly effective when gamma-tACS was applied to the peak of the theta frequency waveform as opposed to the trough. As theta-gamma coupling within the M1 is said to arise spontaneously when GABA activity is blocked [35], a possible explanation is that gamma frequency synchronisation with theta oscillatory activity may improve learning through the mediation of GABAergic activity which may contribute to neural plasticity and subsequent motor learning. However, further study on a range of gamma frequency couplings may clarify the effectiveness of theta-gamma coupling in motor learning. Given its use in coupling, theta-tACS is also becoming considered as a possible modulating frequency for motor learning. Indeed in a recent study [36], theta tACS in a tapping task, with montage located over the cerebellum, resulted in decreased movement regularity, and was concluded to be the result of mediated change in cerebellar output to the M1, consistent with dual-site delivery mentioned above. It should be noted however, that much of the present work on theta-tACS focuses on cerebellar output and inhibition [37], and so at present there does not appear sufficient application of theta-tACS to motor learning explicitly, which we highlight as an area warranting future research.
Although some weak effects of gamma-tACS on motor learning is evidenced from above, much of the evidence is conflicting and further study is required to form a stronger argument for the effectiveness of gamma-tACS on motor learning. Particularly, there is considerable differences in outcome when delivering low or high gamma in either single or dual site montages, likely reflecting the complexity of processes that occur during motor learning, but also indicating the importance of network dynamics (i.e., frequency coupling and dual site connectivity), which have already been shown to be predictive in tACS responsivity [38].
Conclusion and future directions
A summary of the experimental works is provided in Table 1, in which we compare results in the context of task, stimulus settings, montage, and the proposed mechanisms. There is substantial variation in results, largely owing to the task, the placement of stimulation, and the stimulation parameters. We also illustrate activation of brain regions during motor tasks, produced by neurosynth’s [39] automated meta-analysis of 2565 studies relating to the term ‘motor’ (Fig. 1). Here we also estimate placement across the studies mentioned, to illustrate montage in reference to the regions most commonly associated with the term. Inherent to all studies, are many additional parameters including stimulation duration, blinding, and even individual anatomical differences, all of which are factors which many researchers are aware of, and which may indeed contribute to the variability in results.
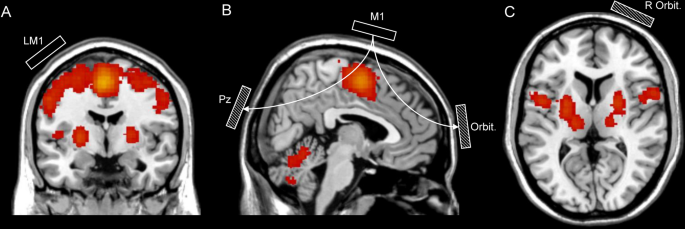
Meta-analysis of regional activation association with the term ‘motor’ from A coronal, B sagittal, and C transverse views. Activation is false discovery rate (FDR) corrected expected at 0.01. Rectangles indicate stimulation (clear) and reference (dashed) electrode placements in the motor cortex (M1), at electrode Pz, and above the frontal orbital (orbit.)
Our current impression of the works discussed here therefore leads to the following conclusions:
Alpha
-
1.
Alpha stimulation to the motor cortex during motor learning improves outcomes, likely modulated by motor cortex excitability changes.
-
2.
Alpha stimulation following motor learning is less cohesive, and appears dependent on the studied population (i.e., age). Indeed age is a very relevant topic in stimulation research, and we speculate likely synergizes further with pre-existing age differences on motor learning performance [40].
Beta
-
1.
Both concurrent and post-task beta stimulation appear to improve learning and retrieval, however most of the evidence is SRTT-based.
-
2.
Beta stimulation is theorized to modulate cortical excitability, however (like alpha) may be dependent on endogenous frequency.
Gamma
-
1.
High gamma (i.e., 70–75 Hz) appears more effective than slow gamma (i.e., 40 Hz) when delivering stimulation concurrent with task.
-
2.
Gamma was more often implicated in cross-frequency coupled effects.
-
3.
Interference with motor retention was most often theorized as the mechanism of negative effect.
The overview presented here is by no means an exhaustive review nor meta-analysis, however consistent findings in the papers discussed suggest that each stimulation frequency has unique mechanisms when modulating (positively or negatively) motor learning and its respective processes. Indeed the exact mechanism of tACS in its varying frequencies remains elusive, and the development of novel hypotheses such as the peripheral nerve mechanism [41] further complicate the interpretation of these works, but offer new avenues to hypothesize the mechanism behind modulation of motor learning. Here we provide some recommendations based on our impression of the works, which fall namely into three major themes: parameters, delivery timing, and montage. We propose that, firstly, more methodological or hypothesis-driven evidence is needed when selecting stimulation frequency and intensity, especially given the variability within the studies discussed here, which ranged from 400 to 2000 μA. This is especially poignant considering work which has put stimulation intensity levels into question [42]. Secondly, the timing of stimulation (whether it is delivered during the task or after the task), is an important consideration especially given that motor learning processes are known to span both online and offline stages [43, 44]. Lastly, the montage used during stimulation is notably important, and depending on whether single or dual site montages are used, different mechanisms could be considered to be at play. Thus, not only is theoretical justification needed for placement, but individual differences (addressed via personalized montages) may also be important in the progression of the field [45]. Evidence has even suggested that intensity-specific effects could be explained by network dynamics [38], highlighting the need for individualized approaches. Indeed the ‘one-tACS-fits-all’ approach could be considered out-dated in the coming years, given that adjacent pharmacological and interventionist fields have already begun to personalize the treatment of individuals, whether it be dose adjustment, or machine learning algorithm. Currently meta-analyses on the effect of tACS on motor learning found a significant positive effect (Z = 3.68, p = 0.0002) across four studies investigating a range of tACS frequencies [46], and another meta-analysis on the effect of beta-tACS on corticospinal excitability found small-to-medium effects on motor evoked potential amplitude [47]. Thus, at present, there appears a small-to-medium effect of tACS in motor learning, which we speculate may actually be limited by the continuation of generic rather than personalized stimulation parameters.
Overall, our recommendation for the continued research of tACS in motor learning culminates into a necessity to provide clear theoretical aims a priori, and rather than maintaining a status-quo of stimulation parameters, justify the selections with scientific and empirically driven reasoning. Approaches which consider individual differences will also be paramount when developing further clinical implementation, particularly given that degree and manifestation of clinical deficits will likely influence treatment efficacy.
No comments:
Post a Comment