If your doctor and therapists can't use this to identify and correct your walking imperfections, then you need to find better ones.
Perturbation-based estimation of within-stride cycle metabolic cost
Journal of NeuroEngineering and Rehabilitation volume 21, Article number: 131 (2024)
Abstract
Metabolic cost greatly impacts trade-offs within a variety of human movements. Standard respiratory measurements only obtain the mean cost of a movement cycle, preventing understanding of the contributions of different phases in, for example, walking. We present a method that estimates the within-stride cost of walking by leveraging measurements under different force perturbations. The method reproduces time series with greater consistency (r = 0.55 and 0.80 in two datasets) than previous model-based estimations (r = 0.29). This perturbation-based method reveals how the cost of push-off (10%) is much smaller than would be expected from positive mechanical work (~ 70%). This work elucidates the costliest phases during walking, offering new targets for assistive devices and rehabilitation strategies.
Introduction
Metabolic cost is a critical measure used to characterize movement behavior [1,2,3]. Healthy walkers naturally adopt an energetically optimal stride cycle, for example, by walking with a step length [4] and knee flexion angle [5] that minimizes metabolic cost. Pathologies like stroke and cerebral palsy alter patients’ walking stride resulting in increases to metabolic cost by 60 to 300% [6, 7]. Such increases in metabolic cost correlate to drastic reductions in people’s mobility and overall quality of life [8, 9]. If we understand how stride cycle phases contribute to metabolic cost, therapies and devices may be better optimized to improve mobility (Fig. 1A).
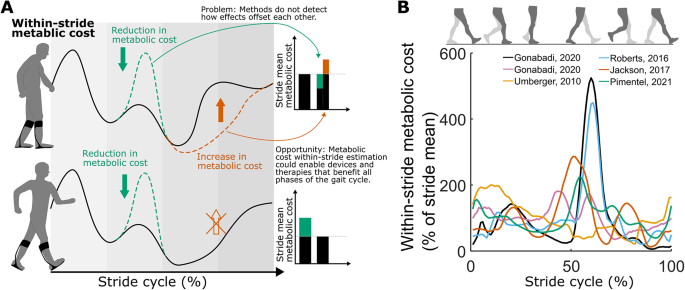
Motivation. (A). Limitation of assessing stride-mean metabolic cost using breath-by-breath measurements. The upper figure illustrates an intervention resulting in a cost reduction (depicted in green) during push-off and a cost increase (depicted in brown) during swing. The stride-mean metabolic cost (displayed in bars) does not enable differentiation of these effects. The lower section of the figure illustrates how comprehending the costs associated with various phases could facilitate the enhancement of interventions. (B). Limited consistency between estimations of within-stride metabolic cost using model-based methods. The mean correlation between estimations is 0.29 (95% confidence interval (CI) = 0.03–0.43) [16, 19, 38, 41, 42]
Measurements of metabolic cost are too slow to detect the contributions of different stride phases. Current methods to calculate energy from oxidative reactions include measuring respiratory CO2 production by ingesting water with a radioisotope (‘doubly labelled water method’), measuring oxidative heat production using a chamber (‘direct calorimetry’), and measuring O2 consumption from respiration (‘indirect calorimetry’) [10]. Indirect calorimetry is the fastest and most commonly used method for measuring metabolic cost during locomotion; however, it still requires averaging several minutes of breaths to be reliable [11,12,13]. A typical walking stride lasts about one second meaning current methods can only measure the mean metabolic cost following a bout of steady-state walking. Experiments that approximated the cost of the swing phase by recording cyclical leg swinging [14] and by measuring blood flow from injected microspheres in animals that are then sacrificed [15] suggest that the stride-mean metabolic cost does not necessarily represent the contributions of individual phases (‘within-stride metabolic cost’).
Several model-based methods of estimating within-stride metabolic cost have been proposed but remain inconclusive. Umberger developed a set of equations to estimate metabolic cost from muscle parameters and used this to produce the first estimation of within-stride metabolic cost from a forward simulation of walking [16]. Other groups used EMG-driven simulations [17] or equations based on joint kinetics instead of muscle parameters [18]. However, when comparing those methods to each other, their estimations of within-stride metabolic cost are relatively inconsistent (Pearson correlation: r = 0.29, n = 6 estimations, Fig. 1B) [19]. Currently, there is no way to validate these model-based estimations for within-stride metabolic cost since measurements from indirect calorimetry only obtain a stride mean. This motivates the development of an alternative method to estimate within-stride metabolic cost that is supported by indirect validation approaches.
We hypothesized that applying a set of perturbations creates a set of instances of the behavior where the differences in the time series between each perturbed instance can be attributed to the different magnitudes and timings of the applied perturbation. By applying perturbations repeatedly to a specific part of the gait cycle for several minutes, we can induce changes in the stride-mean metabolic cost as well as in the biomechanical time series (e.g., kinematics, kinetics, and muscle activations) [19,20,21]. We postulated the variation across the set of perturbed walking strides would be representative of the fluctuations in metabolic cost within the stride cycle so long as the set contained a large number of different perturbations. If true, this would enable a method to extract key features of within-stride metabolic cost. Our approach is inspired by prior studies that utilized ankle perturbations to assess time series of joint impedance during the stance phase [22, 23] as well as studies that used elastic bands and added mass to estimate the cost of stance and swing phases [24, 25]. To the best of our knowledge, using of a perturbation-based approach for estimating within-stride metabolic cost time series is novel.
Using this concept, extraction of within-stride behaviour from a collection of perturbed instances, we developed an alternative method to estimate within-stride metabolic cost that we refer to as our ‘perturbation-based method’. Our method estimates within-stride metabolic cost using measurements from a set of perturbed walking strides. We then evaluated our method’s ability to consistently reproduce model-based estimates of within-stride metabolic cost.
More at link.
No comments:
Post a Comment