What fucking laziness. 'Discuss' NOT PROVIDE PROTCOLS TO RECOVER! We'll never get anywhere in stroke with mentors and senior researchers that allow this crapola to go on. My god, we need to fire a lot of people.
Potential Immunotherapeutic Targets on Myeloid Cells for Neurovascular Repair After Ischemic Stroke
 Ziyu Zhu1†,
Ziyu Zhu1†,  Li Zheng1†,
Li Zheng1†,  Yan Li1,
Yan Li1,  Tingting Huang1, Yu-Chieh Chao1, Lijun Pan2, Hui Zhu1,
Tingting Huang1, Yu-Chieh Chao1, Lijun Pan2, Hui Zhu1,  Yanhua Zhao1,
Yanhua Zhao1,  Weifeng Yu1* and
Weifeng Yu1* and  Peiying Li1*
Peiying Li1*- 1Department of Anesthesiology, Renji Hospital, School of Medicine, Shanghai Jiaotong University, Shanghai, China
- 2Department of Radiology, Renji Hospital, School of Medicine, Shanghai Jiaotong University, Shanghai, China
Neurological deficits and cognitive dysfunctions caused
by acute ischemic stroke pose enormous burden to the stroke families and
the communities. Restoration of the normal function of the
neurovascular unit following ischemic stroke is critical for improving
neurological recovery and cognitive functions after stroke. Recent
evidence suggests that the myeloid cells including both the resident
microglia and infiltrating monocytes/macrophages and neutrophils are
highly plastic in response to the environmental cues. They intimately
interact with multiple components of the neurovascular unit in response
to the alarmins, danger associated pattern molecules (DAMPs) and other
signals released from the ischemic brain. The aim of this review is to
discuss the reciprocal interactions between the myeloid cells and the
ischemic neurovascular unit during the late repair phase of cerebral
ischemic stroke. We also summarize potential immunotherapeutic targets
on myeloid cells and new therapeutic approaches targeting myeloid cells,
such as cell transplantation, mitochondrial dynamic and extracellular
vesicles-based therapy et al to enhance neurovascular repair for better
stroke recovery.
Introduction
The neurovascular unit (NVU) is composed by neurons,
endothelial cells, pericytes, smooth muscle cells, astrocytes, microglia
and extracellular matrix components. It maintains brain homeostasis of
the brain (Banks et al., 2018; Morrison and Filosa, 2019).
All the components of NVU interact with each other to maintain the
neuronal milieu that is required for proper neuronal functioning by
balancing energy, preserving the integrity of the blood brain barrier
(BBB), releasing neurotrophic factors, uptake and recycling of
neurotransmitters and et al. (Halassa et al., 2007; Charveriat et al., 2017).
In response to cerebral ischemic stroke, the integrity of the BBB
compromises leading to leakage of harmful blood components into the CNS,
immune cell infiltration, and aberrant transport and clearance of
molecules. All these changes contribute to the dysfunction of the NVU
which is associated with long term neurological impairments (Fisher and Saver, 2015; Villaseñor et al., 2017).
The reconstruction of NVU and the remodeling of neuronal circuitry
turns out as an essential player in long-term neurological recovery
after stroke (Gurer et al., 2009; Lake et al., 2017).
However, the repair of NVU is a complex process involving clearing of
neuronal debris, neurogenesis, angiogenesis, establishing the new
neuronal circuitry, controlling neuroinflammation and et al. (Lake et al., 2017; Zhao Z. et al., 2017), thus it remains as a big challenge to promote the NVU repair after ischemic brain injury.
After ischemic stroke, both glial cells and peripheral
immune cells can be activated by a variety of mechanisms, including
neuronal “help me” signals, like lipocalin-2 (Xing et al., 2014; Blochet et al., 2018), interleukin 4 (IL-4) (Zhao et al., 2015),
and a complex mixture of extracellular proteins functioning as damage
associated molecular patterns (DAMPs), such as high-mobility group box 1
(HMGB1), hypoxia-inducible factor 1α (HIF-1α), S100B and et al. (Chamorro et al., 2012; Fu et al., 2015; Li et al., 2018b).
Notably, myeloid immune cells are gaining increasing attention due to
their unique function in anti-inflammation, clearing cellular debris and
promoting neuronal plasticity (Iadecola and Anrather, 2011; Shichita et al., 2014; Fu et al., 2015; Zhao S. C. et al., 2017; Wang X. et al., 2018).
Myeloid cells are blood cells that arise from a large
heterogeneous multipotent stem cell population, the hematopoietic stem
cells (HSC) lineage (Kondo et al., 1997; Akashi et al., 2000)
which resides in the bone marrow and possess the ability to give rise
to diverse cell types in the immune system and the blood, including
granulocytes, monocytes, erythrocytes and platelets (Paul et al., 2015).
Some myeloid populations can develop directly from yolk sac progenitors
without apparent bone marrow intermediates, such as tissue resident
macrophages (Kondo et al., 1997; Akashi et al., 2000; Shibata and Suzuki, 2017).
Although peripheral monocyte-derived macrophages and microglia can both
be developed from the same yolk sac progenitors, they should not be
considered as one cellular population in the injured brain (Greenhalgh et al., 2018).
After injury, monocyte-derived macrophages can enter the brain and
directly communicate with microglia and suppress the microglia-mediated
phagocytosis and inflammation after spinal cord injury (Greenhalgh et al., 2018).
In the central nervous system (CNS), the main tissue resident
macrophages including perivascular macrophages (PVM), meningeal
macrophages (MM), and choroid plexus macrophages (CPM) (Henning et al., 2009) and microglia migrate into the CNS during early neural development depending on transcription factor Pu.1 and Irf8 (Hanisch and Kettenmann, 2007; Kierdorf et al., 2013).
In the recent decades, microglia and
monocytes/macrophages are intensively studied in the context of cerebral
ischemic brain injury (Hu et al., 2012; Garcia-Culebras et al., 2018),
with compelling evidence emerging, thus we mainly focused on these
cells in this review. After cerebral ischemia, both residential
microglia and macrophages can be activated within 24 h and the
infiltration of monocyte-derived macrophages and activation of microglia
peaks within 3–4 days after stroke onset (Lalancette-Hebert et al., 2007; Gelderblom et al., 2009; Wattananit et al., 2016).
Polymorphonuclear neutrophil granulocytes (PMNs) are another myeloid
cell population that can infiltrate into the ischemic brain and affect
ischemic brain injury (Frijns and Kappelle, 2002).
These innate immune cells play important roles in determining the
ischemic infarcts. For example, PMNs release matrix metallopeptidase 9
(MMP-9) which degrades the BBB and exacerbates ischemic brain injury (Li et al., 2013a) and tissue-plasminogen activator related hemorrhagic transformation (Mao et al., 2017).
Selective microglia elimination disturbed neuronal calcium responses,
increased calcium overload and increased the incidence of spreading
depolarization, thus significantly increased the infarct size by 60% (Szalay et al., 2016).
Depletion of monocyte/macrophages in cerebral ischemic stroke reduced
hemorrhagic infarct transformation or reduced brain injury in
monocyte/macrophage depleted stroke mice (Gliem et al., 2012; Ma et al., 2016). However, there’s also a recent study showed no impact on stroke outcome with monocyte/macrophages depleted (Schmidt et al., 2017).
Recent researches have highlighted that the impact of
these myeloid cells on the ischemic brain injury largely depend on
different phenotypes at different stages of stroke (Kurisu et al., 2018).
As it’s well-known that microglia and macrophages can be divided into
different phenotypes depending on their functions on the inflammatory
responses (Mantovani et al., 2013; Guruswamy and ElAli, 2017).
Classically-activated (M1) microglia/macrophages usually secret
pro-inflammatory cytokines, such as inducible nitric oxide synthase
(iNOS), tumor necrosis factor α (TNFα), interleukin 23 (IL-23),
interleukin 1β (IL-1β), interleukin 12 (IL-12) and et al. (Mills et al., 2000; Guruswamy and ElAli, 2017; Li et al., 2017),
while alternatively activated (M2) microglia/macrophages are
characterized by their production of anti-inflammatory cytokines, such
as interleukin 10 (IL-10) and transforming growth factor β (TGF-β) (Gordon and Taylor, 2005; Guruswamy and ElAli, 2017) and they can be polarized with IL-4 via a signal transducer and activator of transcription 6 (STAT6)-dependent pathway (Stein et al., 1992; Gao et al., 2015). Recent studies further identified more specific subpopulation of M2 microglia/macrophages, such as M2a, M2b, M2c, and Mox (Hu et al., 2015). Importantly, the phenotypes and functions of these cells are highly dynamic after ischemic injury (Tsuyama et al., 2018).
Likewise, PMNs may also have different phenotypes in different stages
after ischemic stroke, a pro-inflammatory so called N1 and an
anti-inflammatory N2 phenotype (Fridlender et al., 2009).
However, the terminology of microglial/macrophage polarization was questioned and believed to hinder research progress (Ransohoff, 2016). There are far more phenotypes of microglia/macrophages than M1/M2, such as amyotrophic lateral sclerosis specific microglia (Chiu et al., 2013), phagocytic monocyte-derived macrophages (Yamasaki et al., 2014), immunosuppressive CD11c+ microglia and et al. (Kan et al., 2015). There are also cases that microglia/macrophage may concurrently display both pro- and anti-inflammatory phenotypes (Morganti et al., 2016).
In addition, transcriptomics and genomic studies reveal that the
function of microglia are versatile beyond immune responses, such as
synaptic modulation and neurotrophic support (Wes et al., 2016).
Single-cell RNA-sequencing gene profiling showed that macrophages in
traumatic brain injury are not comprised of distinctly polarized cell
subsets, instead, these macrophages are uniquely and broadly activated (Kim et al., 2016). Therefore, it is not rigorous to classify microglia/macrophages imprudently into M1/M2 classifications (Ransohoff, 2016).
Recently, compelling evidence is suggesting the role of
the distinct “healing” myeloid cells during the restoration of NVU after
stroke, including phagocytosis of damaged neuronal debris, promoting
neurogenesis and angiogenesis, re-establishment of the neuronal
circuitry, improving white matter repair and et al. during different
phases after stroke (Figure 1).
FIGURE 1
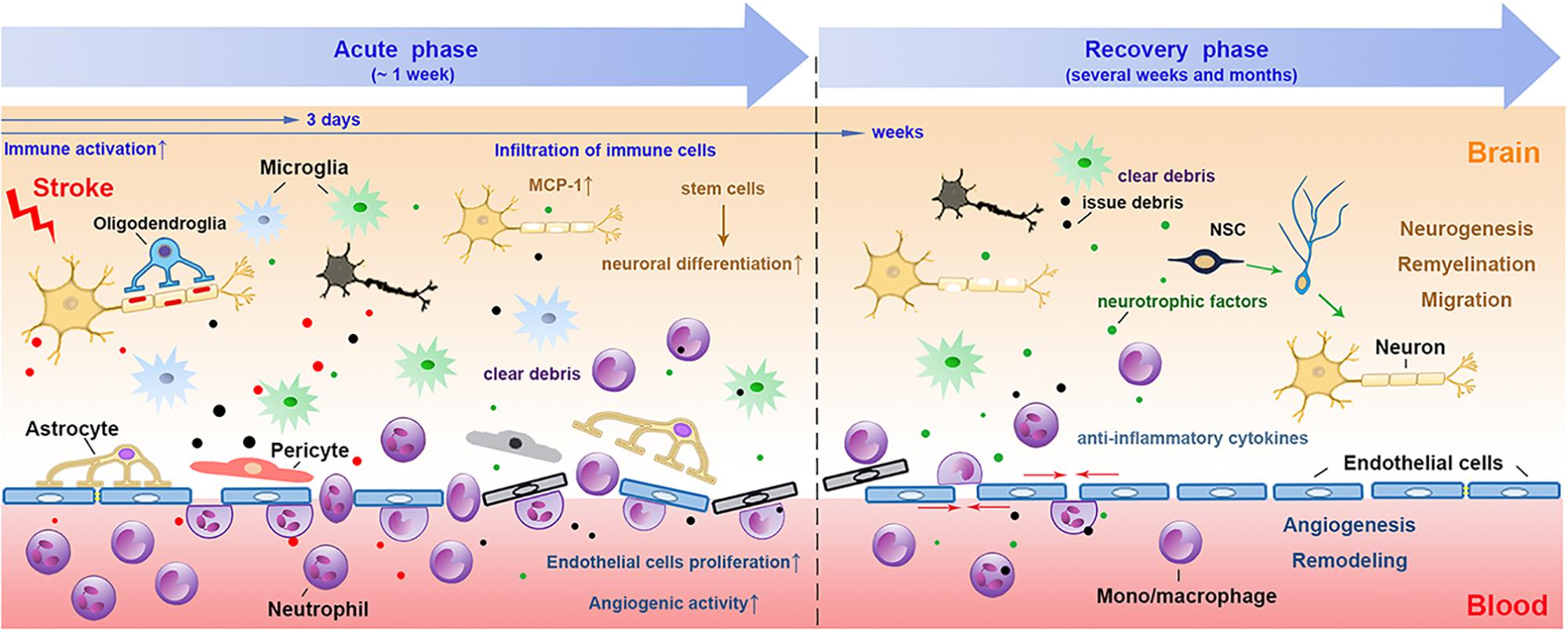 Figure 1. The involvement of myeloid immune cells
during the NVU repair after cerebral ischemic stroke. In the acute phase
of ischemic stroke, the damaged associated signals from the ischemic
brain activate resident and peripheral myeloid immune cells, such as
microglia, peripheral mono/macrophages and neutrophils. These cells
penetrate into the CNS through the disrupted BBB from 1 day after
stroke, peaking at 2–3 days and may last for several weeks. The
phagocytic function of myeloid cells enables them to remove tissue
debris in both acute and repair stages. During the recovery phase after
stroke, myeloid cells with strong plasticity could differentiate into
different phenotypes and release several neurovascular nutritional
factors, which may enhance neurovascular regeneration and remodeling up
to weeks and months after stroke. NVU, neurovascular unit; CNS, central
nervous system; BBB, blood brain barrier; NSC, neural stem cells.
Figure 1. The involvement of myeloid immune cells
during the NVU repair after cerebral ischemic stroke. In the acute phase
of ischemic stroke, the damaged associated signals from the ischemic
brain activate resident and peripheral myeloid immune cells, such as
microglia, peripheral mono/macrophages and neutrophils. These cells
penetrate into the CNS through the disrupted BBB from 1 day after
stroke, peaking at 2–3 days and may last for several weeks. The
phagocytic function of myeloid cells enables them to remove tissue
debris in both acute and repair stages. During the recovery phase after
stroke, myeloid cells with strong plasticity could differentiate into
different phenotypes and release several neurovascular nutritional
factors, which may enhance neurovascular regeneration and remodeling up
to weeks and months after stroke. NVU, neurovascular unit; CNS, central
nervous system; BBB, blood brain barrier; NSC, neural stem cells.
In this review, we will summarize
the distinct functions of different subsets of myeloid cells during the
“repair” of NVU discuss the identified mechanisms underlying their
restoration of the NVU after stroke.
No comments:
Post a Comment