I'm sure your competent? doctor already has protocols on heat and cold shock proteins for your recovery. Oh no, that's not the case!
In Vivo Visualization and Quantification of Brain Heat Shock Protein 90 with [11C]HSP990 in Healthy Aging and Neurodegeneration
Visual Abstract

Abstract
Heat shock protein 90 (Hsp90) is essential for maintaining cellular proteostasis and may play an important role in the development of neurodegenerative proteinopathies. Therefore, we aimed to develop an Hsp90-specific PET brain tracer to quantify Hsp90 expression in the brain in vivo in order to explore its potential as a biomarker for neurodegenerative disease characterization and to support Hsp90-targeted drug development. Methods: We developed the radiosynthesis of (R)-2-amino-7-(4-fluoro-2-(6-(methoxy-11C)pyridin-2-yl)phenyl)-4-methyl-7,8-dihydropyrido[4,3-d]pyrimidin-5(6H)-one, [11C]HSP990, and validated the tracer using in vitro autoradiography, in vitro brain homogenate saturation binding, ex vivo biodistribution, and in vivo PET imaging in rodent models of Alzheimer disease (AD) and Parkinson disease versus healthy age-matched and young controls. Human brain samples from AD patients and healthy subjects were included in our in vitro binding studies. A nonhuman primate PET brain study with arterial blood sampling was conducted under baseline and blocking conditions. Results: In vitro and in vivo [11C]HSP990 studies in rodents and a nonhuman primate revealed saturable Hsp90 binding pools in natural killer lymphocytes, bone marrow, and notably the brain, where the highest binding was observed, particularly in gray matter. Blocking studies indicated that saturable Hsp90 in natural killer lymphocytes considerably influences the pharmacokinetics of Hsp90-targeting probes, which is critical for Hsp90 drug development. In vitro [3H]HSP990 brain homogenate saturation binding assays suggested that the tracer binds a distinct subfraction of the total Hsp90 pool, which is significantly diminished in both rodent and human AD brain tissue compared with age-matched controls. In vivo PET imaging confirmed reduced [11C]HSP990 brain binding on aging and an even stronger decrease in AD mice, suggesting that Hsp90 depletion may impair protein quality control and accelerate proteinopathies. Conclusion: [11C]HSP990 is a promising Hsp90-specific tracer and reveals strong Hsp90 binding in the brain. Uniformly reduced tracer binding was observed in AD brain tissue compared with age-matched controls. [11C]HSP990 holds potential as a biomarker for neurodegenerative disease characterization and progression, and it may aid in patient stratification and therapy monitoring. Human [11C]HSP990 PET neuroimaging studies are under way to investigate whether these findings translate to humans.
Heat shock protein 90 (Hsp90), present in 4 isoforms, is a key chaperone in the protein quality control system, maintaining cellular proteostasis by stabilizing, folding and refolding, and regulating many client proteins (1). Accordingly, Hsp90 function is implicated in diseases associated with proteotoxic stress, such as neurodegenerative disorders and cancer (2,3). The role of Hsp90 in neurodegenerative diseases, including Alzheimer disease (AD), Parkinson disease (PD), amyotrophic lateral sclerosis, and Huntington disease, remains controversial in the literature and has been linked to both protective functions and pathologic roles.
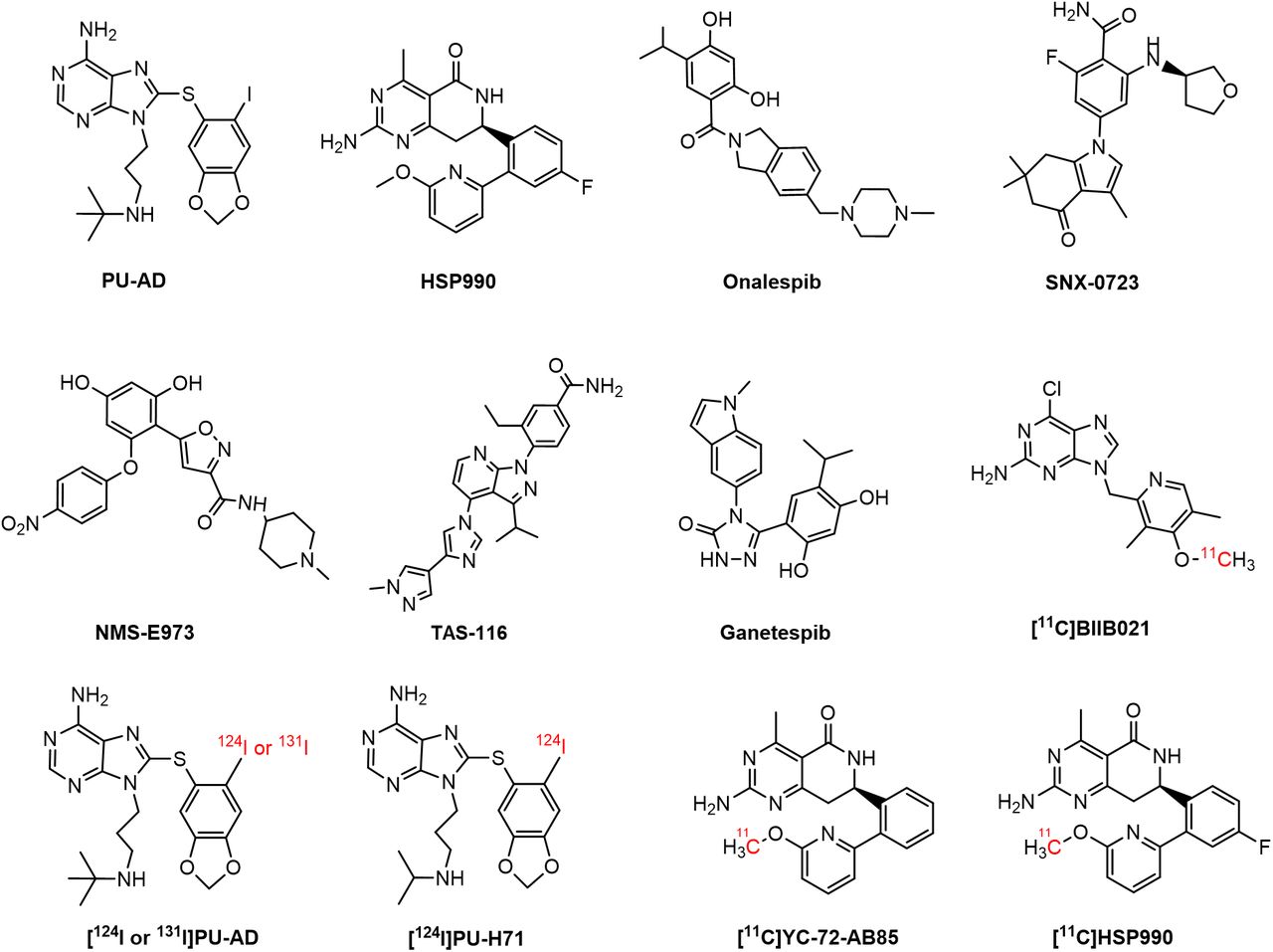
Proteins involved in aggregation, such as β-amyloid, phosphorylated tau, and α-synuclein (αSyn), are reported Hsp90 clients (4–6). Some studies have shown that upregulated Hsp90 colocalized with these aggregates in a neurodegenerative brain, potentially contributing to disease pathology by exacerbating aggregate accumulation and hindering misfolded protein degradation (2,6). This has sparked interest in Hsp90 inhibitors, to suppress its aberrant neuronal activity, as potential treatments for neurodegenerative disorders (7). Conversely, reduced Hsp90 levels have been linked to neuronal cell death. In this context, induction of the heat shock response through Hsp90 inhibition has been explored to upregulate chaperone function, thereby reducing protein aggregation and supporting cytoprotection (1,7–11). For instance, (S)-2-fluoro-6-((tetrahydrofuran-3-yl)amino)-4-(3,6,6-trimethyl-4-oxo-4,5,6,7-tetrahydro-1H-indol-1-yl)benzamide) (SNX-0723) and 9-(3-(tert-butylamino)propyl)-8-((6-iodobenzo[d][1,3]dioxol-5-yl)thio)-9H-purin-6-amine (PU-AD) (Fig. 1) have shown promise in preclinical studies by preventing αSyn oligomerization and rescuing striatal dopamine levels in a PD rat model and by inducing the degradation of misfolded proteins and restoring memory in an AD mouse model, respectively (2,12). PU-AD was evaluated in clinical trials for AD, amyotrophic lateral sclerosis, and glioblastoma, but these studies were withdrawn or terminated and results have yet to be published (13–15). The compound (R)-2-amino-7-(4-fluoro-2-(6-methoxypyridin-2-yl)phenyl)-4-methyl-7,8-dihydropyrido[4,3-d]pyrimidin-5(6H)-one (HSP990) (Fig. 1) has also shown therapeutic potential, improving cognitive function in AD mouse models and reducing huntingtin aggregation in Huntington disease, but exhibited neurotoxicity in a phase I trial for solid tumors (16–18).

{kind=link}
{kind=link}
Chemical structure of Hsp90 inhibitors and PET brain tracers.
The potential clinical efficacy of Hsp90 inhibitors depends on their ability to exert therapeutic effects at safe doses. Accurate measurement of Hsp90 occupancy in the brain using PET can significantly contribute to determining optimal dosing regimens and avoiding toxic effects (19). Furthermore, Hsp90 PET brain imaging may provide insights into the role of Hsp90 in neurologic diseases.
Only a few Hsp90-targeting PET probes that can permeate the blood–brain barrier have been developed so far (2,20,21). [124I]PU-AD (Fig. 1) showed higher hippocampal retention in human AD brain than in controls on static delayed (3 h after injection) PET. However, absolute brain [124I]PU-AD concentrations were very low, and 124I produces low-quality PET images and exposes patients to high radiation because of its long half-life (2,22). More recently, 6-chloro-9-((4-(methoxy-11C)-3,5-dimethyl-2-pyridinyl)methyl]-9H-purin-2-amine ([11C]BIIB021) (Fig. 1) was developed, exhibiting Hsp90-specific binding in the rat brain but showing the presence of brain radiometabolites, which may complicate PET image quantification (21). Our group previously developed (R)-2-amino-7-(2-(6-(methoxy-11C)pyridin-2-yl)phenyl)-4-methyl-7,8-dihydropyrido[4,3-d]pyrimidin-5(6H)-one ([11C]YC-72-AB85) (Fig. 1), demonstrating reversible Hsp90-specific brain binding in healthy rodents and a nonhuman primate (NHP) (20), but it has not been explored in neurodegeneration. Given that HSP990 has already been used in a clinical trial, this study sought to develop and evaluate [11C]HSP990 (Fig. 1) as a Hsp90 PET tracer to elucidate saturable Hsp90 binding and its role in health and neurodegenerative disease and to advance clinical translation.
More at link.
No comments:
Post a Comment