Use the labels in the right column to find what you want. Or you can go thru them one by one, there are only 33,385 posts. Searching is done in the search box in upper left corner. I blog on anything to do with stroke. DO NOT DO ANYTHING SUGGESTED HERE AS I AM NOT MEDICALLY TRAINED, YOUR DOCTOR IS, LISTEN TO THEM. BUT I BET THEY DON'T KNOW HOW TO GET YOU 100% RECOVERED. I DON'T EITHER BUT HAVE PLENTY OF QUESTIONS FOR YOUR DOCTOR TO ANSWER.
Changing stroke rehab and research worldwide now.Time is Brain!trillions and trillions of neuronsthatDIEeach day because there areNOeffective hyperacute therapies besides tPA(only 12% effective). I have 523 posts on hyperacute therapy, enough for researchers to spend decades proving them out. These are my personal ideas and blog on stroke rehabilitation and stroke research. Do not attempt any of these without checking with your medical provider. Unless you join me in agitating, when you need these therapies they won't be there.
What this blog is for:
My blog is not to help survivors recover, it is to have the 10 million yearly stroke survivors light fires underneath their doctors, stroke hospitals and stroke researchers to get stroke solved. 100% recovery. The stroke medical world is completely failing at that goal, they don't even have it as a goal. Shortly after getting out of the hospital and getting NO information on the process or protocols of stroke rehabilitation and recovery I started searching on the internet and found that no other survivor received useful information. This is an attempt to cover all stroke rehabilitation information that should be readily available to survivors so they can talk with informed knowledge to their medical staff. It lays out what needs to be done to get stroke survivors closer to 100% recovery. It's quite disgusting that this information is not available from every stroke association and doctors group.
Sunday, July 19, 2020
Time evolution of frontal plane dynamic balance during locomotor transitions of altered anticipation and complexity
This is incredibly important for you to have delineated in a protocol from your doctor. YOUR DOCTOR'S RESPONSIBILITY! Your doctor will somehow have to extrapolate this from healthy and young to stroke survivors, likely not healthy and not young. Still YOUR DOCTOR'S RESPONSIBILITY! Don't let her run away from that responsibility.
Locomotor
transitions between different ambulatory tasks are essential activities
of daily life. During these transitions, biomechanics are affected by
various factors such as anticipation, movement direction, and task
complexity. These factors are thought to influence the neuromotor
regulation of dynamic balance, which can be quantified using whole-body
angular momentum (H). However, the specific effects of these
factors on balance during transitions are not well understood. The
ability to regulate dynamic balance in the presence of these contextual
factors is especially important in the frontal plane, as it is usually
challenging to maintain walking balance in the frontal plane for
individuals with neuromuscular impairments. The purpose of this study
was to apportion their effects on the time evolution of frontal plane
dynamic balance during locomotor transitions of healthy, unimpaired
individuals.
Methods
Five
healthy young subjects performed 10 separate types of transitions with
discrete combinations of factors including complexity (straight walking,
cuts, combined cut/stair ascent), cut style (crossover, sidestep), and
anticipation (anticipated and unanticipated). A three-way analysis of
variance (ANOVA) was used to compare the maxima, minima, and average
rates of change of frontal-plane H among all transitions.
Results
Before transition, within anticipated state peak value of H increased 307% in crossover style relative to sidestep style (p < 0.0001).
During Transition Phase, within unanticipated state the magnitudes of
average rate of change and peak value increased 70 and 46% in sidestep
style compared to crossover style (p < 0.0001 and p = 0.0003).
Within sidestep style, they increased in unanticipated state relative
to anticipated state. Later in Correction Phase, within both
anticipation states peak value of H increased 41 and 75% in cut/stairs transitions relative to cuts (p = 0.010 and p < 0.0001). For cut/stairs transitions, peak value of H increased 45% in unanticipated state compared to anticipated state (p = 0.0001).
Conclusions
These
results underlined the detrimental effects of unanticipated state and
task complexity on dynamic balance during walking transitions. These
findings imply increased demand of neuromuscular system and functional
deficits of individuals with neuromuscular disorders(like stroke!) during these tasks.
In addition, cutting style influenced frontal plane dynamic balance
before transition and in response to unanticipated direction change.
Collectively, these results may help identify impaired balance control
of fall-prone individuals and inform interventions targeting specific
destabilizing scenarios.
Background
Humans
are frequently required to make cognitive decisions and respond to
external stimuli during walking in uncontrolled “real world”
environments. These cognitive factors can lead to locomotor transitions
that may involve cuts (changing direction), moving from level to uneven
terrain, or complex combinations of these tasks. The ability to perform
these transitions is crucial for activities of daily living, as cuts
alone compose up to 50% of everyday movements [1]. Cutting movements are well-researched in the context of sports movements involving jogging or running [2,3,4,5]. However, much of the literature regarding slower-paced walking tasks have focused on steady-state walking [6,7,8,9], and thus locomotor transitions are not as well understood in the context of typical daily activities.
Steady-state
walking exhibits “orbital stability”, with each step deviating only
slightly from the kinematics of the previous step [10].
In contrast, transitions between locomotion modes are not periodic, and
balance must be maintained while responding to entirely new task
demands. There are two distinct styles of cutting movements, each with
its own distinct medial/lateral biomechanics during walking [11] and running [12, 13]:
a crossover (rotating the trailing, swing leg toward the leading,
implanted leg) or sidestep (rotating the trailing, swing leg away from
the leading, implanted leg). Among several stepping maneuvers related to
these two styles, young and old individuals were more likely to select a
sidestep maneuver to maintain balance in response to walk-in-place
lateral perturbations [14].
While a sport-like agility test indicated the similar preference of
sidestep style, researchers also claimed that sidestep maneuver may
place greater injury risk on the young female athlete relative to
crossover style [13]. Although different cut styles were related to balance regulation during walk-in-place task [14],
it is unclear how healthy adults maintain dynamic balance with each
maneuver during walking cut transitions. Most of previous research on
walking cuts primarily targeted at understanding knee injury mechanisms [15, 16] and joint kinetics [17],
rather than investigating dynamic balance. Moreover, locomotor
transitions may be of movement complexities that could further affect
human biomechanics during walking. For instance, joint power generation
was increased in young adults during transitions of increased complexity
involving both level changes and obstacle avoidance [18].
Transitions from level ground to stair ascent require larger hip and
knee joint moments relative to level walking and thus are
biomechanically challenging [19, 20].
Combined transitions involving both a cut and switching from level
walking to stair ascent are likely even more challenging, but this type
of complex transition has not been thoroughly investigated.
Locomotor
transitions can also be influenced by anticipation, which is a
cognitive rather than environment factor. During locomotion, the nervous
system maintains an “internal model” of the dynamics of the body, and
uses this model in a feedforward sense to coordinate neural control of
movement in preparation for anticipated motions [21, 22].
Unanticipated tasks interrupt this locomotor planning and can therefore
be challenging, especially for individuals with deficits in feedback
neuromuscular control, such as impaired proprioception [23].
For example, the biomechanics of unanticipated cuts may lead to knee
loading mechanics that increase risk of knee ligament injury [15].
In response to unanticipated walk-in-place lateral perturbations, the
young and elderly implemented different stepping maneuvers to maintain
balance but suffered from high frequent collisions of limbs during
stepping [14].
However, the majority of studies that have investigated anticipatory
adjustments during locomotor transitions have focused primarily on joint
mechanics. For example, anticipatory changes of center-of-mass
kinematics, joint angles, and EMG were found before transitioning from
level-ground walking to stair ascent [24].
It is not well investigated how dynamic balance is modified during
unanticipated locomotor transitions. Thus, it remains unclear how the
specific contextual factors of cut style, task complexity and
anticipation affect regulation of dynamic balance during locomotor
transitions.
One metric for assessing dynamic balance during walking is whole-body angular momentum (H), which is tightly regulated by unimpaired individuals during level-ground walking [8]. Regulation of H is achieved primarily through muscle force generation [25, 26]. H is also a valuable quantity to study balance because it relates to the net external moment (Mexternal) about the body center of mass (COM) by the equation H˙=∑Mexternal
. The external moment on body is the cross product of the external moment arm and the ground reaction force (GRF). Thus, H
is directly related to the human whole-body dynamics and is not as
reliant on simplifying kinetic assumptions of inverted pendulum model as
other commonly used measures of dynamic balance, such as margin of
stability [27].
Frontal-plane H
in particular is useful for identifying altered dynamic balance control
in individuals with a variety of neuromuscular impairments. For
example, in individuals post-stroke the magnitude of change in
frontal-plane H during stance of the paretic leg is correlated with lower (worse) Dynamic Gait Index and Berg Balance Scale scores [28]. Although the elderly may have unique strategies performing movement tasks [29, 30], elderly individuals with vestibular balance impairment also have increased frontal-plane H during gait compared to age-matched peers without vestibular dysfunction [31]. The range of H
in people with unilateral transtibial amputation is larger during
prosthetic leg stance compared to able-bodied subjects across several
different walking speeds [32]. During stair ascent, the range of frontal-plane H
is greater compared to level-ground walking in able-bodied individuals,
and is associated with altered GRFs and external moment arms during
stair ascent walking compared to level walking [33].
The unique demands of stair ascent may be particularly challenging for
individuals with neuromuscular impairments. For example, people with
transtibial amputation also have a greater range of frontal-plane H during stair-ascent compared to level-ground walking [34]. However, this increased range of H
and the associated changes in GRFs and external moment arms may be more
difficult to achieve due to reduced proprioception and control in the
prosthesis compared to a biological leg. Furthermore, the transition
from level-ground walking to stair ascent may be more challenging than
steady-state stair ascent. Thus, it is important to understand
regulation of dynamic balance during complex transitions that may pose a
risk to people with neuromuscular impairments. However, dynamic balance
(i.e., regulation of H) before and during these complex
and challenging transitions, particularly when they are unanticipated,
is not yet well understood in unimpaired individuals.
Therefore,
the purpose of this study was to apportion the effects of task
anticipation, cutting style, and complexity on the time evolution of
(i.e. time-varying) frontal-plane dynamic balance during locomotor
transitions of young, healthy, unimpaired individuals based on H. We hypothesized that the peak values of frontal-plane H
would be larger during unanticipated transitions of increased
complexity (i.e., combined cut/stair-ascent). We expected this because
steady-state stair ascent has a larger range of H compared to
level-ground walking, and we expected the interruption of neural
planning during an unanticipated transition to stairs to further
increase the peak values of H. We also hypothesized that able-bodied individuals would have higher average rate of change of frontal-plane H during unanticipated transitions of increased complexity. The average rate of change of H
is equal to the average net external moment about the body COM, and
thus correcting for errors in unanticipated transitions was expected to
increase the required net external moment in the frontal plane.
Methods
Subjects and protocol
Five
young healthy unimpaired individuals (4 females, 1 male) with an
average age of 27.7 (SD = 2.8) years, mass of 52.6 (SD = 2.8) kilograms
and height of 1.68 (SD = 0.06) meters participated in this study. All
participants were free of any known history of neurological or
orthopedic disorders or lower extremity injury prior to the
participation in this study. All participants provided written informed
consent to participate in the experimental protocol that was approved by
the Institutional Review Board. The lab setup consisted of an
over-ground straight-line walkway, a level-ground cutting (45°)
direction to the right, and a mobile staircase at 45° to the left for
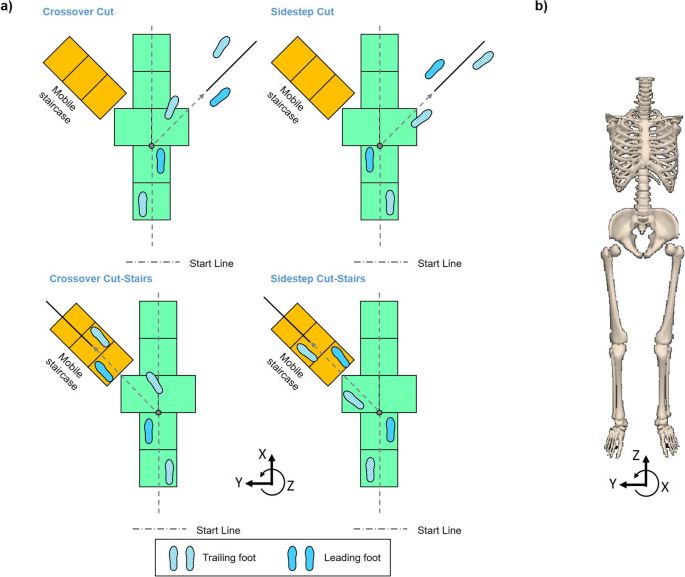
combined cut/stair-ascent (Fig. 1a).
Each participant performed straight-line walking, crossover cut,
sidestep cut, crossover cut/stair-ascent, and sidestep cut/stair-ascent
(Fig. 1b)
under anticipated and unanticipated conditions. First, each participant
completed 10 baseline straight walking trials, followed by 20
anticipated cut trials in a block (5 of each style and complexity)
randomized order. Subsequently, each participant performed 30
unanticipated trials in a fully randomized order, including 10
unanticipated straight walking and 20 unanticipated transition trials (5
of each style and complexity). Short breaks were provided between
sessions. In baseline straight walking, participants were asked to start
with their preferred legs and walk in their normal walking speeds. In
anticipated cut trials, each subject was asked to start with their left
leg for the first 2 blocks and right leg for the last 2 blocks.
Participants were asked to “walk-cut” or “walk-stair” for each block
subsequently. In unanticipated trials, subjects were asked to start with
their left leg for the first block (15 trials) and right leg for the
second block. A randomized auditory cue of “stair”, “cut”, or “walk” was
given at the initiation of single-leg support of the leading leg (i.e.,
the toe-off of the trailing leg), approximately one-half step preceding
a visible transition point (Fig. 1).
The number of each type of auditory cue was controlled to be equal
(5 ± 1 each). The start points were at least 2 steps away from the
transition point.
Fig. 1
a
Diagram depicting experiment setup and a subject preforming crossover
cut (left top) and sidestep cut to stair ascent transitions (right
bottom) using the left (trailing) leg, sidestep cut (right top) and
crossover cut to stair ascent transitions (left bottom) using the right
(trailing) leg. Auditory cue in unanticipated transitions was given at
the initiation of single-leg support of the leading leg (first trailing
leg toe-off). Grey dot on the walkway indicates the transition point. b Diagram depicting the 8-segment human body model and the direction of frontal-plane angular momentum
A
10-camera motion capture system (Motion Lab Systems, Baton Rouge, LA,
USA) operating at 120 Hz was used to track 42 reflective markers placed
bilaterally on the trunk, pelvis, thighs, shanks and feet. An 8-segment
model of each subject (torso, pelvis, thighs, shanks, and feet) was
constructed based on a static trial. Biomechanical data were processed
with Visual3D (C-Motion, Germantown, MD, USA), and 3D motion capture
data were filtered using a low-pass Butterworth filter with cut-off
frequency of 6 Hz. H about the whole-body center-of-mass was calculated in Visual3D as
H⇀=∑8i=1[Iiω⇀+(r⇀i−r⇀COM)×mi(v⇀i−v⇀COM)]
where Ii, ω⇀i
are the moment of inertia tensor and angular velocity, respectively, of the i th segment about the body’s COM in the lab frame; r⇀i and v⇀i are the position and velocity, respectively, of the i th segment’s COM in the lab frame; r⇀COM and v⇀COM
are the position and velocity, respectively, of the body’s COM; mi is the mass of the i th segment. H was normalized by body mass and height of each subject. The direction of H in this study was aligned with the lab reference frame. Positive frontal-plane H indicates rotational momentum toward the “leading leg”, while negative H indicates momentum away from the “leading leg” (Fig. 1b). We defined the leading leg as the implanted leg (stance leg) during turning transition (Fig. 1a).
We analyzed consecutive maxima (P1, P2) and minima (N1, N2) of frontal-plane H,
as well as the average rate of change between each maxima and minima
(P1-N1, N1-P2, P2-N2) during two consecutive strides, from the first
heel strike of the trailing leg to the third heel strike (Fig. 1a). We sub-divided the entire transition movement into four phases (Fig. 1a):
Preparatory, Transition, Correction, and Completion that were defined
by gait events in each leg. The Preparatory Phase occurred before the
cue, starting at the first trailing leg heel strike and ending at
toe-off of the trailing leg (occurrence of the cue). The Preparatory
Phase encompassed positive peak P1. The Transition Phase began with the
cue and lasted throughout leading leg stance, ending with leading leg
toe-off. Transition Phase comprised negative peak N1, and the average
rate of change between P1-N1. Adjustment for errors in the transition
occurred during Correction Phase, which started at leading leg toe off,
lasted throughout trailing leg stance, and ended with the next trailing
leg toe off. Correction Phase included positive peak P2, as well as the
average rates of change between N1-P2. The final phase was Completion
Phase, in which the person reached a new surface to finish the
transition task. Completion Phase was defined as the final leading leg
stance, ending with the final trailing leg heel strike, and comprised
negative peak N2 and the average rates of change between P2-N2.
Statistics
The
Shapiro-Wilk test was performed to check the normality assumption of
the data. Then to determine the effects of anticipation (anticipated,
unanticipated), cutting style (crossover, sidestep), and complexity
(cut, combined cut/stair ascent) on dynamic balance during each phase of
transition, a three-way analysis of variance (ANOVA) was used to
compare the maxima, minima, and average rates of change of frontal-plane
H among all transitions. When the ANOVA indicated significant
main or interaction effects (α = 0.05), post hoc comparisons were
performed to test for significant differences using Bonferroni’s
correction in MATLAB (The MathWorks Inc., Natick, MA, USA). Partial eta
squared (ηp2) was used to calculate effect size
for statistically significant results. Small, medium and large effect
sizes were indicated by ηp2 values greater than 0.01, 0.06 and 0.14, respectively [35].
We then compared the peak values and average rates of change for
anticipated and unanticipated straight-line walk using t-tests
(α = 0.05), and no significant difference was found. Finally, we
compared the peak values and average rates of change of frontal-plane H in transition with straight-line walk using t-tests (α = 0.05).
Results
We analyzed the consecutive peak values of frontal-plane H, as well as the average rate of change between each peak during two consecutive strides (Fig. 2). We observed time lags of the peak H
between different cut styles. Positive peak P1 was found at the first
leading leg heel-strike for all walking tasks. While negative peak N1
was at the tailing leg heel-strike for sidestep style transitions and
straight walk, it was found around the leading leg toe-off for crossover
style transitions. Positive peak P2 was around leading leg toe-off for
sidestep style transitions, while it was around leading leg heel-strike
for crossover style transitions and straight walk. Negative peak N2 was
found near leading leg heel-strike for sidestep style transitions,
trailing leg toe-off for crossover styles, and trailing leg heel-strike
for straight walk.
Fig. 2
Time-varying frontal-plane group averaged H during two consecutive strides of each transition condition.
Black lines represent for anticipated and unanticipated straight-line
walking, while color lines represent for transitions. Solid lines
indicate anticipated conditions, while dash lines indicate unanticipated
conditions. Vertical lines represent chronological events of the first
toe-off of the trailing leg (occurrence of the cue), the first leading
leg toe-off, and the second trailing leg toe off
During
the Preparatory Phase, P1 had significant anticipation and cutting
style main effects, as well as a significant anticipation by cutting
style interaction effect (Table 1).
P1 was significantly different in anticipated transitions compared to
straight walk, while the unanticipated values were not different
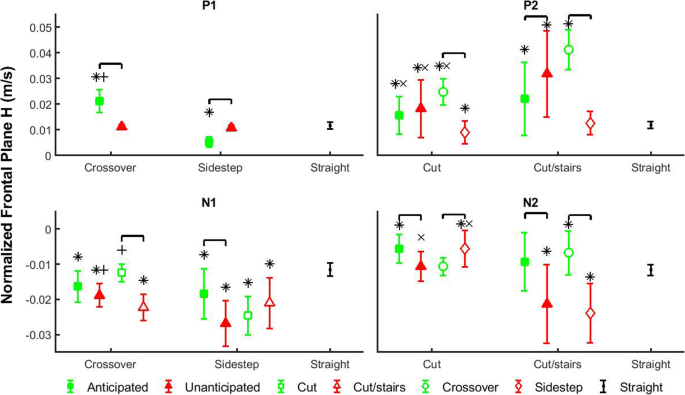
relative to straight walk (Fig. 3). Within anticipated state, P1 increased 307% in crossover style relative to sidestep style (p < 0.0001; Table 2). Furthermore, P1 was larger in crossover styles, but smaller in sidestep style compared to straight walk.
Average and standard deviation of the peak values (P1, N1, P2, N2) of frontal-plane whole-body angular momentum (H)
in each transition phase. Comparisons were performed among anticipation
states (filled shapes), cutting styles (unfilled circles and diamonds)
and complexity tasks (unfilled squares and triangles). Green filled
squares indicate anticipated states, and red filled triangles indicate
unanticipated states. Green unfilled squares indicate cuts, and red
unfilled triangles indicate cut/stairs transitions. Green unfilled
circles indicate crossover styles, and red diamond indicate sidestep
styles. Each marker above represents an interaction scenario of two
fixed factors, and consists of different levels of the other fixed
factor. Comparisons were also performed between each transition scenario
and straight walk. Black dots indicate straight walk in both
anticipation states. Brackets indicate significant differences between
two transition scenarios. Significant differences between each
transition scenario and straight walking are indicated by ‘*’. ‘+’
indicates significant differences between crossover and sidestep cut
styles within the same anticipation states, and ‘×’ indicates
significant differences between cut and cut/stairs within the same
anticipation states or the same cut styles
During
the Transition Phase, the average rate of change between P1-N1 had
significant anticipation and cutting style main effects, as well as
anticipation by cutting style and cutting style by complexity
interaction effects. The negative peak N1 had significant anticipation,
cutting style, and complexity main effects, as well as anticipation by
cutting style, and cutting style by complexity interaction effects.
Within unanticipated states, the magnitudes of P1-N1 and N1 increased 70
and 46%, respectively, in sidestep style compared to crossover style (p < 0.0001 and p = 0.0003), while they are not different within anticipated states (Fig. 3, Fig. 4).
For crossover style, the magnitudes of both P1-N1 and N1 increased 32
and 78%, respectively, in more complex cut/stairs transitions relative
to cuts (p = 0.037 and p < 0.0001). However, for
sidestep style the magnitudes of P1-N1 and N1 increased 56 and 43%,
respectively, in unanticipated states compared to anticipated states (p < 0.0001 and p = 0.0006).
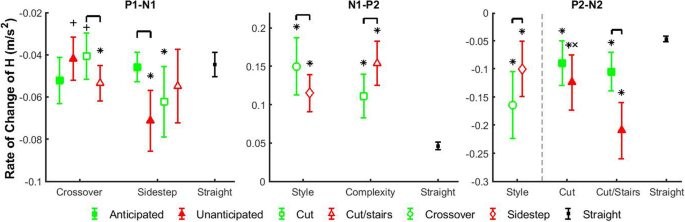
Fig. 4
Average
and standard deviation of the average rate of change (P1-N1, N1-P2,
P2-N2) of frontal-plane whole-body angular momentum (H) in
different transition phases. Comparisons were performed among
anticipation states (filled shapes), cutting styles (unfilled circles
and diamonds) and complexity tasks (unfilled squares and triangles).
Green filled squares indicate anticipated states, and red filled
triangles indicate unanticipated states. Green unfilled squares indicate
cuts, and red unfilled triangles indicate cut/stairs transitions. Green
unfilled circles indicate crossover styles, and red diamond indicate
sidestep styles. Each marker above represents a walking condition of a
main factors or an interaction scenario of two main factors, and
consists of different levels of interactions or main factor,
respectively. Comparisons were also performed between each transition
scenario and straight walk. Black dots indicate straight walk in both
anticipation states. Brackets indicate significant differences between
two transition scenarios. Significant differences between each
transition scenario and straight walking are indicated by ‘*’. ‘+’
indicates significant differences between crossover and sidestep cut
styles within the same anticipation states, and ‘×’ indicates
significant differences between cut and cut/stairs within the same
anticipation states or the same cut styles
During
the Correction Phase, the average rate of change between N1-P2 had
significant main effects of cutting style and complexity. N1-P2
increased 30% in crossover style relative to sidestep style (p < 0.0001), and increased 38% in cut/stairs transitions compared to cuts (p < 0.0001).
All main effects of positive peak P2 were significant, and significant
anticipation by complexity and cutting style by complexity interaction
effects were also found. Within both anticipated and unanticipated
states, P2 increased 41 and 75%, respectively, in cut/stairs transitions
relative to cuts (p = 0.010 and p < 0.0001). For more complex cut/stairs transitions, P2 increased 45% in unanticipated state compared to anticipated state (p = 0.0001). P2 increased 66% in cut/stairs transitions relative to cuts for crossover style (p < 0.0001),
while it was not different for sidestep style. Finally, for both cut
and cut/stairs tasks, P2 increased 176 and 227%, respectively, in
crossover style compared to sidestep style (p < 0.010 and p < 0.0001).
During
the Completion Phase, all main effects of the average rate of change
P2-N2 were significant, and the anticipation by complexity interaction
effect was also significant. In general, the magnitude of P2-N2 was 64%
larger in crossover transitions relative to sidestep transitions (p < 0.0001).
For more complex transitions, the magnitude of P2-N2 was 100% greater
in unanticipated state compared to anticipated state (p < 0.0001).
Negative peak N2 had significant anticipation, cutting style, and
complexity main effects, as well as anticipation by complexity and
cutting style by complexity interaction effects. For both cuts and
cut/stairs transitions, the magnitude of N2 increased 91 and 127%,
respectively, in unanticipated state relative to anticipated state (p < 0.021 and p < 0.0001). For sidestep style, the magnitude of N2 was 327% larger in cut/stairs transitions compared to cuts (p < 0.0001).
In addition, within unanticipated states, both the magnitudes of P2-N2
and N2 increased 69 and 99%, respectively, in cut/stairs transitions
relative to cuts (p < 0.0001 and p < 0.0001).
Discussion
Increased task complexity and unanticipated state pose challenges on dynamic balance regulation
We investigated the time-varying frontal-plane dynamic balance based on whole-body angular momentum (H)
during transitions of altered anticipation, cut style and complexity in
healthy young adults. Our first hypothesis was that the peak values of
frontal-plane H would be larger during unanticipated transitions
of increased complexity, such as unanticipated cut/stairs. This
hypothesis was supported. During Correction Phase when participants
transitioned from level ground to stair-walking (Fig. 1a),
positive peak angular momentum P2 was larger in cut/stairs transitions
relative to cuts, regardless of anticipation state (Fig. 3).
Furthermore, within cut/stairs task, P2 was larger in unanticipated
state compared to anticipated state. While it has been shown that
increased range of frontal-plane H was maintained in steady-state stair ascent compared to straight level walk for healthy subjects [36], our results further suggest that increased H was required for unanticipated cut/stairs transitions. The increased peak value of frontal-plane H
may indicate higher risk of mediolateral fall during unanticipated
complex transitions, which is affected by interrupted neuromuscular task
planning in unanticipated states [15] and enhanced joint moment requirement for stair walking [19, 20].
To further understand the regulation of H, we investigated the average rate of change of H
that can be considered as the average net external moment about the
body’s COM. Our second hypothesis was that the average rate of change of
frontal-plane H would also be largest during unanticipated
transitions of increased complexity. This hypothesis was partially
supported. The magnitude of the average rate of change of H
between N1-P2, during the Correction Phase, was larger in
cut/stair-ascent transitions compared to cuts, while there was no effect
of anticipation state (Fig. 4).
Decreased mediolateral GRFs and vertical moment arms that are main
contributors to the mediolateral net external moment likely explain the
enhanced slope and range of frontal-plane H during early stance of stair ascent relative to level straight walking [36].
Similarly, the increased magnitude of N1-P2 slope indicating larger
mediolateral net external moment may explain the increased positive peak
P2 during stair-ascent transitions. Furthermore, the gluteus medius has
been shown to be the major contributor to maintain frontal-plane
angular momentum by rotating the body toward the ipsilateral leg during
stance phase [26].
It has been also reported that gluteus medius performed similarly in
maintaining mediolateral balance in stair and level walking [37].
Positive angular momentum relates to rotation toward the leading leg,
consistent with the functional direction of gluteus medius in
maintaining dynamic balance. Thus, the increased frontal-plane H
may result from reduced gluteus medius activity during a level-stair
transition. Previous study also suggested that increased frontal-plane
angular momentum during steady-state stair walking may be a necessary
strategy to raise body COM while avoiding a trip [36].
Therefore, complex locomotor transitions from level to stair-ascent
walking may require a different strategy relative to level transitions,
and excessive angular momentum swinging human body toward leading leg
may be needed for dynamic balance. These results may be useful for
assessing risks of balance-challenged populations during complex
locomotor transitions.
Anticipatory changes in dynamic balance are influenced by cut style
Individuals
make cognitive adjustments for their control of dynamic balance before
they approach the transition point, and their strategies depend
exclusively on cut styles according to our results. During Preparation
Phase participants in anticipated states increased the positive peak of H
(P1) for crossover styles, but reduced the positive peak for sidestep
styles compared to straight walking and unanticipated transitions (Fig. 3). Although we did not analyze H before P1, style-specific modifications on anticipatory H were also performed at the first trailing leg heel strike (Time 0) with the same strategies used for P1 (Fig. 2). These adjustments are understandable because the increased H (rotation toward the leading leg) for crossover styles and reduced H
(rotation away from the leading leg) for sidestep styles were the same
as the cut styles. These findings may be a generalized strategy in
prepared human walking cuts. A recent study found that during
anticipated 90-degree walk turns, angular momentum was not affected by
the direction of change as long as individuals use sidestep cut style [38]. Similar conclusions were also made for healthy individuals performing crossover 90-degree cuts to right and left direction [39].
Although these investigations did not compare different cut styles,
their results support our findings that anticipatory change of dynamic
balance is affected by cut style, but not the direction. Furthermore,
these adjustments of whole-body angular momentum may partially result
from preparatory control of trunk angular momentum that had the same
modification strategy as H [40].
Previous study on sidestep cuts also reported that trunk swing is a
strategy assisting in moving body COM toward new walking direction [41].
They found that the trunk displaced opposite from the cutting direction
before turning, and assisted direction change in an inverted pendulum
manner. However, this different trunk strategy was thought to be used in
a late cue (unanticipated) transition, and contribute less in an early
cue (anticipated) condition. Therefore, individuals in anticipated
walking cuts initiated their control of mediolateral dynamic balance in
advance to prepare for the direction change, and the swing of upper body
segment may contribute to this strategy.
These anticipatory adjustments on H
may also have implications on the effects of different transition
factors on walking dynamic balance regulation. Our results showed that
individuals modified their dynamic balance at least one half step before
anticipated transitions. This is expected because studies have shown
gaze fixation on the future foot landing area before at least two steps [42, 43], modifications on gait parameters for two strides before transition to stairs [24], and high rate of successful direction change when individuals were cued two steps ahead [44]. Nonetheless, subjects adjusted H
based on cut styles, not task complexity that requires more
biomechanical changes. Although it could be argued that individuals
prioritize some more closely approaching challenges, they failed to
adjust H for task complexity at least half step before
transitions. This may suggest a priority of cut style over task
complexity (cut/stairs) in the “internal model” of human nervous system
that regulates locomotion and dynamic balance. This priority may be due
to the fear of knee injury during cuts that are associated with
increased breaking forces (anterior/posterior GRF) and quadriceps
activation [12].
Therefore, healthy adults pre-rotated their body to prepare for
incoming cut transitions, which may be an effective strategy to maintain
dynamic balance and avoid injuries during these destabilizing tasks.
Our findings on the anticipatory adjustments of healthy dynamic balance
regulation may provide a baseline to evaluate and improve related
routines in rehabilitation training.
Reactive control of dynamic balance is influenced by cut style
Individuals
make reactive changes to dynamic balance in response to unexpected
auditory cue of transition, and the strategies are different for each
cut style. During sidestep-style transitions, they increased the
magnitudes of P1-N1 and N1 for unanticipated states compared to
anticipated states, but during crossover styles the magnitudes remain
for both anticipation states (Fig. 3, Fig. 4).
Furthermore, within unanticipated states the magnitudes of P1-N1 and N1
were greater in sidestep style compared to crossover style, which may
be due to the unique mechanisms that individuals used for each cut
style. There was a delay of the occurrence of the negative peak N1
during crossover-style transitions compared to straight walking and
sidestep-style transitions (Fig. 2).
It was at the trailing leg heel-strike for sidestep style transitions
and straight walking, but at the subsequent leading leg toe-off for
crossover styles. Moreover, H in crossover-style transitions was
maintained as tightly as straight walking at the trailing leg
heel-strike. With limited response time in unanticipated conditions
(auditory cue at the initiation of leg swing), participants were not
able to change leg swing trajectories rapidly to cross the stance leg,
and an unanticipated gait termination was performed in crossover
transitions. Although angular momentum was tightly regulated,
unanticipated crossover transition with gait termination may still be
challenging for balance-impaired populations [23].
Furthermore, as direction change continued participants in
unanticipated crossover transitions used the leading leg to turn to the
new direction in a sidestep style. However, this is only an initiation
of direction change because H was still in the direction away
from the leading leg, i.e., opposite to the new direction of travel.
This mechanism was similar to the previously reported control strategy
of body COM in the initiation of walking direction change where the
trunk was displaced to the opposite of new direction [41]. However, individuals in unanticipated sidestep transitions may easily swing the trailing leg and trunk [40]
away from the leading leg direction with increased negative momentum.
Thus, in response to unanticipated walking direction change crossover
style may require rapid gait termination and inverted-pendulum-style
trunk motion to initialize its direction change, while sidestep may take
advantage of the momentum during leg swing to be a more effective
maneuver for quicker changing of locomotion direction [12].
Nonetheless, the potential balance challenge in the gait termination of
unanticipated crossover transitions and increased momentum in
unanticipated sidestep transitions may still need to be carefully
considered in rehabilitation training.
Limitations and future considerations
One limitation of our study is that we did not include arms in the model used to calculate H. Although arm swing may contribute to transverse-plane H during treadmill walking [45] and anteroposterior fall recovery [46], evidence have shown that the magnitude of contributions to frontal-plane H from the arms are very small relative to the trunk and legs during normal walking [8] and 90-degree turn [38].
Our statistical results may also be limited by the modest number of
participants. To mitigate this, we collected five trials of each
condition for each subject and analyzed the results objectively. While
we used subject-averaged data for analysis, the effect sizes turned out
to be large (ηp2 > 0.14) for all statistically
significant results. Nonetheless, the results should be interpreted as
initial findings given the modest number of participants. Another
limitation may be that we evaluated H in the lab (inertial) frame compared to recent research on dynamic balance in the body moving reference frame [47]. H
in inertial frame can be directly related to ground reaction force
measurements. We also believe that during walking turns, dynamic balance
in the direction of inertial frame is more endangered, evidenced by
significantly larger peak values compared to H in anatomical medial/lateral direction (not published). Finally, H
in this study was not normalized by walking velocity. Although previous
studies showed different angular momenta with walking speed [9, 32], we wanted to incorporate the velocity information in the single metric H,
which may be part of the strategy that individuals used to maintain
dynamic balance during walking transition, as increased speed of
transition can adversely influence walking stability of both young and
old individuals [48].
Future work is also needed to understand segmental contributions to
angular momentum in each transition task so that specific strategy of
dynamic balance control in locomotor transitions can be apprehended and
targeted for rehabilitation training. Future experiment and analysis on
patients walking during locomotor transitions may also be useful to
fully understand dynamic balance regulation mechanism and improve
rehabilitation training of these populations.
Conclusions
The
results of this study underlined the detrimental effects of
unanticipated states and task complexity on dynamic balance during
walking transitions. These results imply increased demand of
neuromuscular system and functional deficits of individuals with
neuromuscular disorders during these tasks. In addition, cutting style
influenced frontal plane dynamic balance. These healthy young adults
made anticipatory adjustments of dynamic balance before transition based
on cutting style, but not complexity (i.e., terrain). Reactive control
of dynamic balance in response to unanticipated direction change was
also affected by cutting style with specific advantage and challenge to
the locomotion tasks.

No comments:
Post a Comment